Plasmamembran: Struktur og funksjoner av plasmamembran
Les denne artikkelen for å lære om membranstrukturen, transportmodifikasjonen og funksjonene til plasmamembranen!
Plasmamembran:
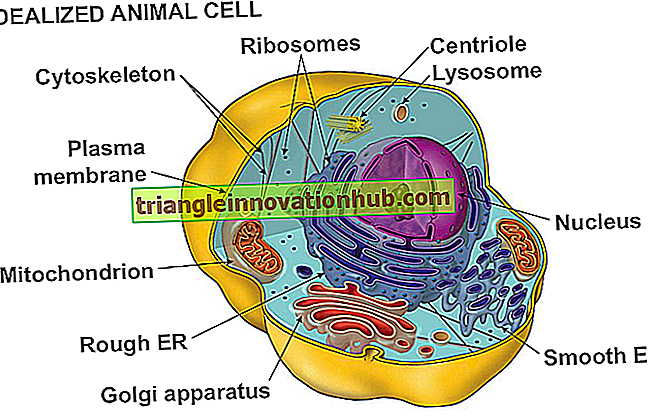
Plasma membran eller plasma-lemma er en bio-membran som forekommer på utsiden av cytoplasma i både prokaryoter og eukaryotiske celler.
Det skiller den cellulære protoplasma fra sitt ytre miljø. Prokaryotiske celler har ikke indre membranpartisjoner. Sistnevnte forekommer i eukaryotiske celler som dekning av flere celleorganeller som kjerne, mitokondrier, plastider, lysosomer, Golgi-legemer, peroksisomer etc.
Biomembraner linjer endoplasmatisk retikulum. De forekommer også på thylakoider i plastids eller cristae inne i mitokondriene. Vacuoles er separatzd fra cytoplasma av en membran som kalles tonoplast. Alle biomembraner er dynamiske i naturen, og viser kontinuerlig endringer i form, størrelse, struktur og funksjon. Plasma membran ble oppdaget av Schwann (1838). Den ble kalt membran av Nageli og Cramer (1855). Membranen ble gitt navnet på plasma lemma av Plowe (1931).
Kjemisk natur av membraner:
Kjemisk består en bio-membran av lipider (20-40%), proteiner (59-75%) og karbohydrater (1-5%). De viktigste lipidene i membranen er fosfolipider (ca. 100 typer), steroler (f.eks. Kolesterol), glykolipider, sfingolipider (f.eks. Sfingomyelin, cerebrosider).
Karbohydrater tilstede i membranen er forgrenede eller ikke-forgrenede oligosakkarider, f.eks. Heksose, fukose, heksoamin, sialinsyre, etc. Proteiner kan være fibrøse eller globulære, strukturelle, bærer, reseptorer eller enzymatiske. Omtrent 30 typer enzymer er registrert i forskjellige biomembraner, f.eks. Fosfataser, ATP-ase esteraser, nukleaser, etc.
Lipidmolekylene er amfetiske eller amfipatiske, det vil si at de har både polare hydrofiliske (vannkjærlige) og ikke-polare hydrofobe (vannavstøtende) ender. Den hydrofile regionen er i form av et hode mens den hydrofobe delen inneholder to haler av fettsyrer.
Hydrofobe haler skjer vanligvis mot midten av membranen. Proteinmolekyler har også både polare og ikke-polare sidekjeder. Vanligvis er deres polare hydrofile koblinger mot ytre side. De ikke-polare eller hydrofobe bindingene holdes enten brettet innvendig eller brukes til å etablere forbindelser med hydrofob del av lipidene. Flere typer modeller har blitt fremsatt for å forklare strukturen til en biomembran. Jo viktigere er Lamellar og Mosaic.
Lamellar Modeller (= Sandwich Modeller):
De er de tidlige molekylære modellene av biomembraner. Ifølge disse modellene antas bio-membraner å ha en stabil lagdelt struktur.
Danielli og Davson Modell:
Den første lamellarmodellen ble foreslått av James Danielli og Hugh Davson i 1935 på grunnlag av deres fysiologiske studier. Ifølge Danielli og Davson inneholder en biomembran fire molekylære lag, to fosfolipider og to av proteiner. Fosfolipider danner et dobbeltlag.
Fosfolipid-dobbeltlaget er dekket på hver side av et lag av hydrerte globulære eller a-proteinmolekyler. De hydrofile polarhoder av fosfolipidmolekylene er rettet mot proteinene. De to holdes sammen av elektrostatiske krefter. De hydrofobe, ikke-polare haler av de to lipidlagene er rettet mot senteret der de holdes sammen av hydrofobe bindinger og van der Waals-krefter.

Robertson Modell:
J. David Robertson (1959) endret modellen av Danielli og Davson ved å foreslå at lipid-dobbeltlaget dekkes på de to flater med utvidede eller (3-proteinmolekyler. En forskjell i proteinene i ytre og indre lag ble også foreslått, f.eks. mukoprotin på ytre side og ikke-mucoid protein på innsiden.
Robertson jobbet på plasmamembranen av røde blodlegemer under elektronmikroskop. Han ga begrepet enhetsmembran som betyr at:
(i) Alle cytoplasmiske membraner har en lignende struktur av tre lag med et elektron-gjennomsiktig fosfolipid-dobbeltlag som er sand-hekset mellom to elektroner tette lag av proteiner,
(ii) Alle biomembraner er enten laget av en enhetsmembran eller et flertall av enhetsmembran. Enhetsmembranen til Robertson kalles også trailaminar membran. Den har en tykkelse på ca. 75 Å med et sentralt lipidlag med 35 A tykk og to perifere proteinlag med 20Aeach. Ifølge Robertson, hvis en membran inneholder mer enn tre lag, eller er tykkere enn 75A, må den være et flertall av enhetsmembran.
Mosaic Modell:
Væskemosaikkmodell. Det er den nyeste modellen av en biomembran som ble foreslått av Singer og Nicolson i 1972.
1. Ifølge denne modellen har membranen ikke en jevn disposisjon av lipider og proteiner, men er i stedet en mosaikk av de to. Videre er membranen ikke solid, men er kvasevæsken.
2. Det postulerer at lipidmolekylene er til stede i et viskøst billag som i lamellarmodellen. Proteinmolekyler forekommer på steder både i og på ytre siden av lipid-dobbeltlag. De interne proteiner kalles inneboende eller integrerte proteiner, mens de ytre er kjent som ekstrinsiske eller perifere proteiner.
Integrert eller inneboende proteiner utgjør 70% av de totale membranproteinene og passerer inn i lipid-dobbeltlaget til forskjellige dybder. Noen av dem løper gjennom lipid-dobbeltlaget. De kalles tunnelproteiner som individuelt eller i en gruppe danner kanaler for passasje av vann og vannløselige stoffer.
3. Proteinene gir den strukturelle og funksjonelle spesifisitet til membranene. Siden lipid-bilaget er kvasifluidt, kan membranproteinene skifte lateralt og derved gi fleksibilitet og dynamikk til membranen.
Mange membranproteiner virker som enzymer, noen av dem oppfører seg som per-meases for letter diffusjon og noen få proteiner fungerer som bærere fordi de aktivt transporterer forskjellige stoffer over membranen. Visse andre proteiner fungerer som reseptorer for hormoner, anerkjenningssentre og antigener. Noen av lipidene på ytre overflaten er kompleksdannet med karbohydrater for å danne glykolipider eller glykoksyx.

Modifikasjoner av cellemembran:
1. Microvilli:
De er finger som evaginasjoner på 0, 6-0, 8 μm lengde og 0, 1 μm diameter som er funnet på den frie overflaten av celler som er involvert i absorpsjon, f.eks. Tarmceller, leverceller, mesothelceller, uriniferøse tubuli. Overflaten som har microvilli kalles strikket kant eller penselgrense.
Microvilli øker overflaten flere ganger. De støttes av en web av mikrofilamenter, actin sammen med myosin, tropomysoin, spektrin, etc. De smale mellomrom mellom mikrovilli deltar i pinocytose.

2. Mesosomer:
De er plasmalemma infoldings funnet i bakterier. En type mesosom er festet internt til nukleoid. Det kreves for nukleoidreplikasjon og celledeling.

3. Junctional komplekser:
De er kontakter mellom tilstøtende celler som i tilfelle av dyrceller er separert med mellomrom på 150-200 Å fylt med vævsvæske. De viktigste er:
(i) Interdigitasjoner:
Det er sammenkobling av fingerlignende membranutviklinger mellom to tilstøtende celler. Interdigitasjoner øker kontaktområdet mellom to celler for utveksling av materialer.
(ii) Intercellulære broer:
Fremskrivninger fra tilstøtende celler gjør kontakt for rask ledning av stimuli.
(iii) Stramme veikryss:
(Zonulae Occludentes, singular-Zonula Occludens). Her blandes plasmamembraner av to tilstøtende celler i en rekke punkter med et nettverk av rygg eller tetningstrenger. Stramme veikryss forekommer i epitel med høy elektrisk motstand, og hvor filtrering skal skje gjennom cellene, for eksempel kapillærer, hjerneceller, samling av nyrer.

(iv) Gap Junctions:
De tilstøtende cellene har protoplasmiske forbindelser gjennom spesielle proteinflasker kalt connexons. Hver konnexon er laget av seks identiske proteinunderenheter rundt en hydrofil kanal.

(v) Plasmodesmata:
De er protoplasmiske broer mellom planteceller som forekommer i områdene av cellevegger eller porer.
(vi) Desmosomer:
(Maculae Adherentes, singular-Macula Adherens). Tilstøtende membraner har skiveformede fortykkelser på ca. 0, 5 (am diameter, et antall tonofibriller (= tonofilamenter) og transmembran-linkere innebygd i tett intercellulært materiale. Desmosomer fungerer som punktsveis og kalles følgelig spot desmosomer. De forekommer i epithelia underkastet å forstyrre.

(vii) Terminalbjelker:
(Belt Desmosomes, Zonulae Adherentes, Singular-Zonula Adherens. Intermediate Junction). Terminalstenger er desmosomer uten tonofibriller. Bånd av fortykning forekommer på den indre overflaten av membranen. Båndene inneholder mikrofilamenter og mellomliggende filamenter.
Funksjoner av cellemembraner:
1. Hovedfunksjonen til cellulære membraner er compartmentalisation. Som plasmamembraner separerer de cellene fra deres ytre miljø. Som organelle belegninger tillater de cellemellene å opprettholde sin identitet, spesifikke indre miljø og funksjonelle individualitet.
2. Membranene tillater strømmen av materialer og informasjon mellom forskjellige organeller av samme celle, så vel som mellom en celle og en annen.
3. Som plasmodesmata og gapforbindelser, gir biomembranene organiske forbindelser mellom tilstøtende celler.
4. Plasma membraner samt andre membraner i organellene har selektiv permeabilitet, det vil si at de tillater at bare utvalgte stoffer passerer innover til utvalgte grader. Membranene er ugjennomtrengelige for andre.
5. Biomembraner har egenskapen til retentivitet, det vil si at de ikke tillater utadgående passering av stoffer som allerede er tillatt oppføring.
6. Plasma membran har spesifikke stoffer på overflaten som fungerer som anerkjenningssentre og vedlegg.
7. Stoffer festet til cellemembranen bestemmer antigen-spesifisitet. Glykophoriner tilstede på overflaten av erytrocytter fungerer som antigen-determinanter. Histokompatibilitetsantigener indikerer om en fremmed celle eller vev skal inkorporeres eller avvises.
8. Cellmembran har reseptorer for visse hormoner. Hormonet kombinerer med sine spesielle reseptorer og endrer enten membranpermeabilitet eller aktiverer enzymadenylatcyklase for å produsere syklisk AMP fra ATP. cAMP utløser deretter et sett med enzymer for å utføre en bestemt funksjon.
9. Membraner har bærerproteiner for aktiv transport.
10. Cellemembraner inneholder enzymer for å utføre viss reaksjon på overflaten, f.eks. ATP-ase (for ATP-syntese og frigjøring av energi fra ATP), fosfataser, esteraser etc.
11. Visse cellemembraner (f.eks. Plasmamembran i bakterier, tylakoidmembraner av kloroplaster, indre mitokondriamembran) har elektrontransportsystemer.
12. Membran infolds brukes til masseinntak av materialer ved endocytose.
Membran Transport:
Passasje av stoffer over biomembraner eller cellemembraner oppnås ved følgende metoder:
A. Transport av vann:
(I) Osmose:
Osmose er diffusjonen av vann eller løsningsmiddelmolekyler gjennom plasmamembran fra lavt osmotisk trykk til høyt osmotisk trykk, dvs. fra høyt vanninnhold til lavt vanninnhold. Plasmemembranen fungerer som en differensialmembran som tillater bevegelse av vannmolekyler inn og ut som holder metabolittene.
B. Transport av joner og små molekyler:
(II) Passiv transport:
Det er en modus for membrantransport der cellen ikke bruker energi eller viser spesiell aktivitet. Transporten er i henhold til konsentrasjonsgradient. Det er av to typer, passiv diffusjon og letter diffusjon.
(a) Passiv diffusjon eller transport over cellemembraner:
Her spiller cellemembranen en passiv rolle i transport av stoffer på tvers av den. Passiv diffusjon kan oppstå enten gjennom lipidmatrise av membranen eller ved hjelp av kanaler.
(i) lipidoppløselige stoffer:
Det ble funnet ut av Overton (1900) at lipidoppløselige stoffer passerer raskt over cellemembranen i henhold til deres konsentrasjonsgradient. Basert på dette funnet, overton foreslått at cellemembraner er laget av lipider.
(ii) Kanal Transport:
Membran har kanaler i form av tunnelproteiner som ikke bærer noen ladning. De tillater vann og oppløselige gasser (CO 2 og O 2 ) å passere i henhold til konsentrasjonsgradienten. Osmose er et eksempel på en slik transport.
Hvis to løsninger med forskjellige konsentrasjoner separeres av en semi-permeabel membran, beveger løsningsmiddelmolekylene seg over membranen fra de mindre konsentrerte til den mer konsentrerte løsningen. Denne prosessen - diffusjonen av løsningsmiddelmolekyler i en region der det er en høyere konsentrasjon av løsemiddel som membranen er impermeabel, kalles osmose.
Filtrering er diffusjon under trykk over en membran som har små porer. Ultrafiltrering skjer under glomerulær filtrering i nyrer. Dialyse er prosessen med å separere små partikler (f.eks. Krystallinske oppløsninger) fra større (f.eks. Kolloider) på grunn av forskjell i diffusjonshastigheten over en membran som har meget små porer.
(b) Tilrettelagt diffusjon:
Det skjer gjennom etaten av spesielle membranproteiner kalt permeaser. Når en slik transportmediert transport er fra et område med større konsentrasjon, er det ikke nødvendig med energi, og prosessen kalles for letter diffusjon. Som et resultat er transporthastigheten stereospesifikk.

Innføring av glukose i blodkropp i røde blodformer er en forenklet diffusjon.
Prosessen med forenklet diffusjon innebærer følgende trinn:
1. Diffusjonsmolekyler kombinerer med de spesifikke bærerproteinmolekyler som danner bærer-proteinkomplekser.
2. Formen til bærerproteinmolekylet endres som respons på diffusjonsmolekylet slik at de membranbundne bærerproteinkompleksene danner kanaler.
3. Formen av bærerproteinmolekylet endres som respons på diffuserende molekyl, slik at molekylet kan krysse plasmamembranen.
4. Når diffusjonsmolekylet har nådd den andre siden, reduserer forandringen i form av bærermolekyl (konformasjonsendring) dens affinitet med det diffuserende molekylet og tillater det å bli frigjort.
5. Etter frigjøringen av diffusjonsmolekylet gjenopptar bærerproteinmolekylet opprinnelig form.
Den lette diffusjonen gjør det mulig for molekyler å krysse den ellers impermeable eller dårlig permeabel membran.
Tilrettelagt diffusjon avviker fra enkel diffusjon i følgende funksjoner:
(i) Tilrettelagt diffusjon er stereospesifikt (enten L- eller D-isomer transporteres).
(ii) Det viser metningskinetikk.
(iii) Tilrettelagt diffusjon krever en bærer for transport over membranen. Bæreproteinmolekylene beveger seg frem og tilbake over membranen ved termisk diffusjon.
(III) Aktiv transport:
Det er oppoverbakke bevegelse av materialer over membranene hvor de oppløste partiklene beveger seg mot deres kjemiske konsentrasjon eller elektrokemisk gradient. Denne transportformen krever energi som tilveiebringes nesten utelukkende ved hydrolyse av ATP.
Aktiv transport skjer i tilfelle både ioner og ikke-elektrolytter, for eksempel saltopptak av planteceller, ioner, glukose og fenolftalein i tilfelle nyretubuli, natrium og kalium i tilfelle av nerveceller etc. Det støttes av ulike bevis:
(a) Absorbsjon reduseres eller stoppes med redusert oksygeninnhold i omgivelsene.
(b) Metabolske hemmere som cyanider hemmer absorpsjon.
(c) Celler akkumulerer ofte salter og andre stoffer mot konsentrasjonsgradienten.
(d) Aktiv transport viser metningskinetikk som er, transporthastigheten øker med økning i oppløsningskonsentrasjon til et maksimum oppnås. Utover denne verdien øker hastigheten til membrantransporten ikke noe som indikerer at den foregår gjennom byrået med spesielle organiske molekyler kalt bærermolekyler, bærerpartikler eller bærerproteiner.
Bærermolekylene er ATP-aser, enzymer som katalyserer hydrolysen av ATP. Den viktigste av disse ATPasene er Na + -K + ATP ase som også er kjent som Na + -K + -pumpen. Det er i tillegg H + -K + ATPaser i mageslimhinnen og nyrene.
Det er et spesielt bærermolekyl for hver oppløst partikkel. Bæreren har sitt bindingssted på to overflater av membranen. De oppløste partiklene kombineres med bæreren for å danne bærer-løsemiddelkompleks. I bundet tilstand gjennomgår bæreren en konformasjonsendring som transporterer oppløsningen til den andre siden av membranen. Energi brukes til å oppnå konformasjonsendringen i transportøren. Det er levert av ATP. I prosessen dephosphoryleres ATP for å danne ADP. Bæreproteiner er av tre typer.
1. Uniport:
De transporterer bare ett stoff.
2. Symptom:
I noen tilfeller krever transport binding av mer enn ett stoff til transportproteinet og stoffene transporteres over membranen sammen. Et eksempel er symporten i tarmslimhinnen som er ansvarlig for samtransporten ved å lette diffusjonen av Na + og glukose fra tarmlumen til slimhindeceller.
3. Antiports:
De bytter ut ett stoff til en annen. Na + -K + ATPase er en typisk antiport.

Mange dyrceller opererer en natrium-kalium utvekslingspumpe ved deres plasmamembran. En lignende protonpumpe opererer i kloroplaster, mitokondrier og bakterier Na + -K + -utvekslingspumpen opererer ved hjelp av enzym ATP-ase som også fungerer som bærermolekyl.

Enzymet hydrolyserer ATP for å frigjøre energi. Energien brukes til å oppnå konformasjonsendringer i transportøren. For hver ATP-molekyl hydrolyseres, pumpes tre Na + -ioner utover og to K + -ioner pumpes innover.
Na + - K + -utvekslingspumpen utfører følgende funksjoner: (i) Opprettholder et positivt potensial på ytre siden av membranen og relativt elektronegativ potensial på innsiden,
(ii) Pumpen skaper et hvilepotensial i nervecellene,
(iii) Pumpen opprettholder vannbalansen i levende celler.
(iv) Det hjelper i dannelse av urin,
(v) Det deltar i utskillelse av salt som hos marine dyr. Havmåler og pingviner drikker sjøvann. De skiller ut overflødig salt gjennom nesekjertlene. Nese saltkjertlene har natrium-kaliumpumpe i plasmamembranen i cellene deres. Na + -ioner pumpes ut aktivt. Klorioner passerer passivt ut. Nesekresjon av de to fuglene har 1, 5-3, 0 ganger mer NaCl-konsentrasjon enn den som er tilstede i blodet.
(vi) Det usekretiserte og umagnetiserte overskudd Na + -ioner som er til stede i det ekstracellulære fluidet har en tendens til å passere tilbake i cellene. Andre stoffer kombineres med natriumioner og passerer innover med dem, for eksempel glukose, aminosyrer i tarmen. Fenomenet kalles sekundær aktiv transport sammenlignet med Na + -K + utvekslingspumpe som kalles primær aktiv transport.

Andre viktige pumper inkluderer Kalsiumpumpe (RBC, muskler), K + pumpe, CP-pumpe, K + -H + -vekselpumper. Den siste forekommer i vaktsceller.
Aktiv transport er et middel for (i) absorpsjon av de fleste næringsstoffer fra tarmen (ii) reabsorpsjon av nyttig materiale fra uriniferrørene (iii) rask og selektiv absorpsjon av næringsstoffer av celler (iv) opprettholde et membranpotensial (v) vedlikehold av hvilepotensial i nerveceller (vi) opprettholder vann og ionisk balanse mellom celler og ekstracellulær væske, (vii) utskillelse av saltkirtler.
C. Transport av faste partikler (Bulk transport):
Bulk transport innover så vel som utover skjer over plasmamembranen ved invaginering og evaginering av membranen. Bulk transport er nyttig i å transportere store molekyler som ville ha problemer med å passere gjennom cellemembranen normalt. Endocytose og eksocytose er de to måtene hvor bulktransport er oppnådd.
(IV) Endocytose er prosessen med å oppsluke store partikler av matstoffer eller av fremmede stoffer. Ifølge stoffets natur kan endocytosen være:
(i) Pinocytose eller celler som drikker er prosessen med inntak av væskemateriale av cellen.
(ii) Mikro pinocytose er pinocytosen av subcellulært eller submikroskopisk nivå.
(iii) Rhophaeocytose er overføringen av små mengder cytoplasma sammen med deres inkludering.
(iv) Fagocytose er oppslukt av partikler av stor størrelse med fast mat eller fast stoff av cellen.
(V) Eksocytose er prosessen med å eksudere sekretærproduktene til utenfor cellenes cytoplasma. Det er også kjent som emeicytose eller celleoppkast. I celler i bukspyttkjertelen beveger de vakuolholdige enzymer seg fra det indre av cytoplasma mot overflaten. Her smelter de sammen med plasmamembranen og tømmer innholdet til utsiden.