Hagfish: Anatomi og fysiologi (Intro, fordøyelsessystem og andre organer)
I denne artikkelen vil vi diskutere om: - 1. Introduksjon til Hagfish 2. Hagfish biologi 3. Eksterne egenskaper 4. Alimentary Canal 5. Excretory System 6. Åndedrettssystem 7. Kardiovaskulærsystemet 8. Reproduktive System.
Innhold:
- Introduksjon til Hagfish
- Biologi av Hagfish
- Eksterne egenskaper av Hagfish
- Alimentary Canal of Hagfish
- Excretory System of Hagfish
- Åndedrettssystem av Hagfish
- Det kardiovaskulære systemet av Hagfish
- Reproduktive System av Hagfish
1. Introduksjon til Hagfish:
Cyclostomata tilhører superklasse Agnatha. Det er representert av levende former som hagfishes og lamprays. De er egentlig ikke fisk. Iktyologer har imidlertid gjerne tatt dem til studier fordi de er akvatiske vertebrater som vanligvis forekommer i forbindelse med ekte fisk. Til tross for deres primitive strukturelle egenskaper, er lampreys og hagfishes begge vellykkede grupper som er mer rikelig enn de fleste innser.
Opprinnelsen og forfeden til denne gruppen er diskutabel og forstås ikke fullt ut. Endoskelettet av disse dyrene består av brusk og derfor blir deres fossile arkiver ikke tilfredsstillende bevart i geologisk rekord.
Men ifølge noen arbeidere har de steget ned fra ostracoderms, en gruppe av gamle utdøde pansrede fisk, på grunn av følgende årsaker:
1. De har et enkelt nesebor i midten av hodet.
2. De har ikke nedre kjeve.
3. De mangler parede finner.
4. Det er ingen benaktig vertebral kolonne.
Klassesyklostomata er delt i to undergrupper:
1. Myxinoidea
2. Petromyzontia
Personene som tilhører Myxinoidea er kjent som hagfishes. Linnaeus klassifiserte dem som ormer. De er representert av omtrent to dusin arter, de vanligste er Myxin, Eptatretus (Bdellostoma) og Paramyxin.
Myxin er bredt fordelt langs kysten av Nord-Europa og Atlanterhavskysten av Nord-Amerika. Noen arter oppstår i kysten av Chili og Japan. Eptatretus stoutii er funnet vidt distribuert langs Pacific Coast av både Nord-og Sør-Amerika, i kystvann i Sør-Afrika og New Zealand.
2. Biologi av Hagfish:
Hagfishes er omfattende havboere og bor vanligvis på sjøbunnen i gravene. De bor ikke bare i gjørmete burger, men lever i døde eller døende fisk og forbruker kjøtt og viskose som bare forlater hud og bein. De finnes bare i lavt vann med lav temperatur (15 ° C), der vannet er salt og kaldt.
Ifølge Jensen (1961-1966) er Eptatretus stoutii beboer den vestlige kontinentalsokkelen i Nord-Amerika fra Alaska til Baja California i dybden, spenner fra så lite som 60 føtter til så mye som 1800 fot.
Hagfishen er svak i naturen og forblir stille i gjørmete burrows, men når det er sulten, beveger det seg raskt med slange, men normalt er bevegelsen sløv eller torpid eller kjedelig. Kroppsfarge er gråbrun til blåaktig brun over, hvitaktig under.
Hagfish har bemerkelsesverdig immunitet mot infeksjoner og sykdommer. De har ikke tymuskjertel, men aggregering av lymfoide celler i velar muskler har vært antydet som primitiv prototype struktur. De produserer heller ikke antistoff.
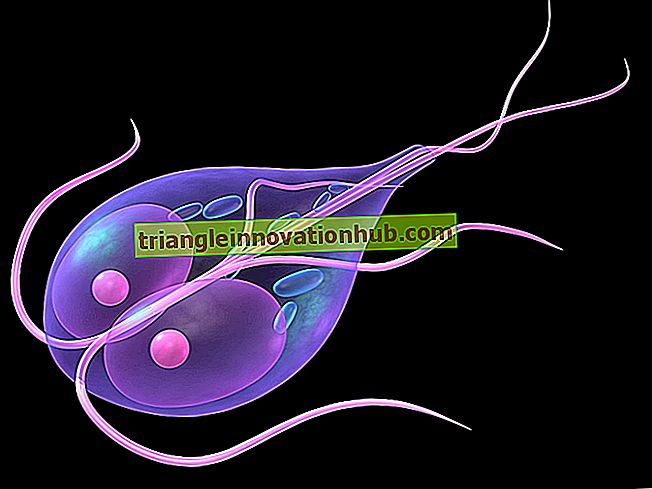
3. Eksterne egenskaper av Hagfish:
Disse kalles ofte som slime ål fordi de avskiller tykk slipperslime i enorme mengder med fantastisk hurtighet. En enkelt hagfish vil fylle en to gallon bøtte med slime blandet med vann i noen sekunder og etter et lite intervall kan gjenta operasjonen enkelt.
Slimmet blir utskilt av små porer på hver side av ventraloverflaten som strekker seg fra snute til spissen av halen. Porene utskiller melkefluid som i kontakt med vann danner trådlignende fast kokong rundt hagfish. Denne strukturen er ekstremt nyttig for å unngå å gripe av noen rovdyr.
Hagfish kommer ut av denne kokongen ved å lage sløyfer og knuter i kroppen som gir hagfish ekstra innflytelse og dermed trekker dyret seg ut av slimmet og unngår chocking av gjær. Hagfishes er bemerkelsesverdig for deres mangel på slående egenskaper. Forreste har den en rund munn, som er forsynt med tre par skiver eller kjøttfulle tentakler (figur 2.1a, b).

De er taktile i naturen, det vil si at de har følelse av berøring. Hodet har et enkelt nesebor, en særegen egenskap av syklostomata. Den har rudimentære øyne, men dyret er helt blindt på grunn av dyphavsadaptasjon.
Imidlertid er de i stand til å skille mellom mørkt og lys på grunn av pigmentet som er tilstede i øyets øyhinne. Noen følsomme områder er også til stede på hodet og rundt cloaca. Det er om en dozon gill åpninger som ligger på hver side på den ventrale overflaten av kroppen.
De ser ut som hvite flekker. I Myxine er det bare ett par. Dorsfinen er fraværende eller udefinert smal og ikke delt. Caudalfinen er litt utviklet. Preanalfin er tilstede.
4. Fôrkanal av Hagfish:
Fordøyelseskanalen til hagfish er et rett rør som består av munn som åpner i halsen, som fører inn i tarmen gjennom svelg, som slutter til anus (figur 2.2a, b, c). Myxins munn er hesteskoformet, omgitt av to par tentakler. Tungen er utstyrt med to rader med små tenner.
Tennene oppstår fra tannplaten. Tunge og sugeapparatet er godt utviklet. Tarmene kan deles inn i forut og hindgut. Nær halsen er det et fremtredende sirkulært muskelbånd som avgrenser disse to regionene. Det er ingen mage i Eptatretus.
I noen arter er magen litt et dilatert kammer. Deres sirkulære muskelbånd forhindrer vannet å komme inn i fordøyelsessystemet, men mens dyret forbruker mat, slapper musklene av bandet og lar maten komme inn i fordøyelsessystemet. Tarmene er utstyrt med rygger som øker tarmens overflate for absorpsjon av ernæring, men spiralventilen er fraværende.
Lever, galdeblære og bukspyttkjertel er ekstrautstyr fordøyelseskanaler og åpne i tarmene. Bukspyttkjertelen av hagfish er delt inn i to helt separate deler, en utskiller fordøyelsessaft, det andre insulin.
Hormoninsulinet regulerer formentlig blodsukkernivået i hagfish som det gjør hos andre vertebrater. Tarmsystemet har dobbel funksjon, på den ene siden er det fordøyelseskan mens det derimot er hemopoietisk, dvs. produserer røde blodlegemer.
5. Ekskresjonssystem av Hagfish:
Nyrene av Myxin er delbare i to deler, pronephros og mesonephros som er segmentert ordnet. Protefrosene beholdes i voksen hagfish som ikke er merket av mesonephros. I Eptatretus er pronephrickanalen tilstede, men Myxin mangler slike kanaler. Det er ingen urinproduksjon av pronephros, men har fagocytiske og hemopoitiske funksjoner.
I Myxin og Eptatretus inneholder pronephros mer enn 2000 ciliated nephrostomes (peritonealtunneler). Mesonephros er funksjonell nyre og består av malpighiske kroppslegemer (Bowmans kapsel), som inneholder glomerulus. Denne delen mangler tubulater som de andre vertebrater.
Mesonephros er hovednyrene og er ansvarlig for å filtrere blodet for utskillelse eller reabsorpsjon i kroppen. Hagfisken kan ikke overleve lenge i vann som enten har en veldig høy eller svært lav konsentrasjon av salter. Det osmotiske trykket i blodet styres av adrenokortiske hormoner.
6. Åndedrettssystem av Hagfish:
Det er et dusin eller så par gillpouches arrangert langs kroppen. De åpner i svelget. Myxine har seks par gill poser mens Eptatretus er utstyrt med 13 til 15 par gill poser.
Hver glidelåse gir ut utløpsrør, ca seks utgangsrør åpner i en felles kanal som åpner utvendig med en enkelt glidelås i Myxine, men i Eptatretus åpnes hver glidelåse separat, derfor er det tretten til femten par ytre glidelåser ( Fig. 2.2a, b, c).



Vannet kommer inn i halsen gjennom nesebor, forut er lukket på grunn av virkningen av bånd av sirkulær muskel og hindrer vannstrømmen i tarmen. Velumet, et par muskelflapper som ligger i halsen, retter vannet inn i glideposer og deretter går vannet ut gjennom gissel ved hjelp av kanaler.
Kanalen som kalles pharyngocutaneous duct er tilstede umiddelbart bak den siste glidelåsen og på venstre side åpner den til ekstern grunnlinje. Det hjelper i fjerning av store partikler fra strupehodet og unngår inngangen til delikate gillpouches. Gullene er forsynt med blodkar og blodstrømmen i motsatt retning til vannstrømmen, og dermed gjør motstrømssystemet (figur 2.3).

Derfor er det raskt utveksling av oksygengass. Slike mekanismer er også til stede i fiskene.
7. Det kardiovaskulære systemet av hagfisk:
Kardiovaskulærsystemet består av blodkar, som sirkulerer blod til og fra alle deler av kroppen, hjertet som holder blodet i bevegelse, og lymfekarene som sirkulerer lymf. Sirkulasjonssystemet er særegent fordi venøs sirkulasjon ikke er forbundet med arteriell side på vanlig måte.
I flertallet av organer er det kapillær seng, og arterielt blod blir tømt i kapillærsengen, men på et sted er det ingen kapillær seng, men arterielt blod åpner i bihulene gjennom hvilke det perkolerer før det kommer inn i venen.
En åpen sirkulasjon av denne typen er vanlig hos hvirvelløse dyr, men sjelden hos vertebrater. Resultatet av en slik sirkulasjon er at blodtrykket på venøs side er svært lavt.
Hovedhjertet av hagfish er kjent som grenhjerte. Bortsett fra grenhjerte (fig. 2.4a, b) er hagfisk særegent fordi den har hjerteorganer, og disse er (a) kaudalt hjerte, (b) portalhjerte, (c) kardinalhjerte. Branchial hjertet ligger stengt for bilobed leveren.
Det avgrenede eller systemiske hjertet til syklostomer ligger i et bruskbeende, som kommuniserer med bukhulen i hagfisk, som Myxine via en peri-kardio peritonealkanal. Grenhjerte består av en sinus venosus, et atrium, en ventrikel og en bulbus arteriosus som ligner på fiskene (figur 2.5).


Kransløpssirkulasjonen er helt fraværende i atrium og ventrikel i grenfibrens hjerte, men koronarfartøy forekommer i veggen av sinus venosus av Eptatretus burgari.
Sinus venosus er et tynt vegget kammer, det venøse blodet fra de ulike kroppsdelene blir mottatt gjennom ducti Cuvieri og andre blodkar. Den høyre ductus Cuvieri er utryddet, så bare venstre kanal av Cuvieri vedvarer i voksen alder.
Sinus venosus består av glatt muskel i veggen og åpner seg for atrium. Sinuatrialåpningen er utstyrt med ventil som hindrer tilbakestrømningen. Atriumet er enkelt og ligger over midtlinjen. Den åpnes i ventrikkel gjennom atrioventrikulær åpning som er bevoktet av atrioventrikulære ventiler. Ventrikken åpnes i bulbus arteriosus.
Myokardiet i atriumet og ventrikken er svampet, musklene deres er ordnet i ledninger eller trabeculae som projiseres inn i lumen i atriumet og ventrikkelen. Trabeculae er mer tett pakket og større i ventrikkelen enn atrium. Bulbulen åpnes i ventral aorta. Det er kort og ikke-kontraktil.
Elektronmikroskopisk struktur av hjertemuskulaturcellene i syklostomene er studert og vurdert av Yamauchi (1980). Den fine strukturen til cyclostomata (lamprey og hagfish) muskel ligner den hos skjelettmuskulaturen i vertebrater.
A- og I-båndene er tydelig skilt. Jeg båndet er lett og har Z-linje, A-båndet er mørkt og leveres av H-bandet. M-linjen halverer H-båndet og N-linjen er tydelig synlig. Ifølge Leak (1969) er N-linjen ubetydelig i hagfish, Myxine glutinosa, men det er ganske fremtredende i Lampetra.
I myxin glutinosa demonstrerte Helle et al (1972) spesifikke myokardiske granulater sammen med de funksjonelle granulene av perifer kobling av sarkoplasmisk retikulum, har en proteinholdig matrisesubstans som sannsynligvis vil være assosiert med kalsium. Kjernen, Golgi-komplekset, multicellulære legemer, ribosomer og glykogenpartikler er tilstede.
Det skal påpekes at det ikke foreligger noen kriterier for å diskriminere, med hensyn til forekomst eller fravær av noen spesielle organeller, hjertemuskelceller mellom sinus venosus, atrium og ventrikel i hjertet av syklostomer.
Epikardiet skiller seg ut i en enkelt kontinuerlig mesothelialcelle og bindevev i både Eptatretus burgeri og Lampetra japonica. Kjellerlaminatet er tydelig. Kjernen og Golgi-komplekset, endoplasmatisk retikulum, ribosomer og pinocytotiske granulater er rikelig.
Endokardiumforingen i hagfisken viser en forholdsvis jevn tykkelse, dvs. 300 nm, som helt er uten fenestre eller intracellulære hull og er utstyrt med overflod av pinocytotiske vesikler. Kjelleren lamina er til stede i hagfish men fraværende i lamprey.
Myokard-cytoplasma inneholder ovoide eller nesten sfæriske granuler i vesikler med en diameter på 100-300 nm karakteriserer cytoplasmaet av sekretoriske intestinale celler (SICs). Disse granulatene inneholder katekolaminer og ligner på chromaffin-celler av adrenalmedulla samt kromaffinlignende celler i hjertet av amfibier, reptiler og pattedyr.
Hepatisk portal Hjerte av Hagfish:
Leverportalven i kurset nær leverlobene blir utvidet og pulserende og er kjent som leverportalveinhjertet. Det er rørformet og grovt T-formet. Den ligger mot nedre høyre side av hagfishen.
Det er mindre og enkelt i struktur og slår med raskere hastighet. Helle, (1972) var av den oppfatning at SIC granulatene (katekolamin) i portalens hjerte av myxin glutinosa er mer elektron-tette enn ventrikkelen til samme organisme.
Hjertet er ikke et neural fordi endokardial bindevev i portalen hjertet av Eptatretus burgeri inneholder nervebunter. Ifølge Carlson (1904) og Fange, (1963), er det den forstørrede delen av sinus supra-intestinalis som er kjent for å pulsere som en pacemaker for portalvein-hjerte.
Caudal Hjerte av Hagfish:
Det kaudale hjertet består av parret utvidelse eller sacs, en på hver side av en median bruskplate (figur 24b). Dens både innløp og uttak er bevoktet av ventiler. Disse sacs samler sinusvæsken fra subkutane bihuler i haleområdene og pumper væsken i efferente kar, som forener for å danne en median kardinalvein.
Disse sacs er ikke kontraktile i seg selv, men fylles og tømmes vekselvis ved virkningen av ekstrinsiske somatiske muskler (musculi cordis caudalis) som løper på utsiden av hver og festes til bruskplater. Det kaudale hjertet er aktivert av nerver. Den har et reflekssenter i ryggmargen.
Ifølge Jensen (1966) vil stimuleringen av hagfishhuden hemme det kaudale hjerteslaget helt. En ren neurogen sammentrekning av muskelen på den ene siden forårsaker bøyning av platen mot den siden som resulterer i en fylling av sacen på den samme side og samtidig tømning av sekken på motsatt side. Den har A- og I-bånd med H, M, N og Z linjer som er typiske for skjelettmuskulaturen.
Kardinalhjerte av Hagfish:
Kardinal hjertet er par sac plassert i hodet regionen. Det har ingen muskler eller maskiner som er egne for å produsere rytmiske impulser; dens pumpevirkning drives av rytmisk sammentrekning av skjelettmuskler som ligger utenfor sac i hodegruppen. Blodet fra dette kardinalhjertet via kardinalårene går til gren- eller portalhjertet.
Sirkulasjon av blod i hagfisk:
Blodet fra hovedgren eller systemisk hjerte pumpes inn i ventral aorta gjennom bulbus arteriosus (figur 2.6).

Fra ventral aorta går blodet til gjellene gjennom avferente grenarterier. Etter rensing oppsamles blodet av efferente greniale arterier og helles i dorsal aorta.
Blodet fra dorsal aorta leveres til fremre og bakre side av kroppen gjennom en serie blodårer, de store blodkarene er karoten arterier, som tilfører blod til kroppens muskel og hud, da blir en gren gitt av for å levere blod til fremre og bakre lobes i leveren, da er det gitt en gren som gir blod til gonader og til nyre og så til slutt til haleområdet.
Blodet går fra arterier til blodårene gjennom kapillærene som er tilstede i sengen i gjellene, tarmene og kroppens muskler. I hodet slippes blod fra arterier ut i store åpne områder som kalles bihuler eller sinusoider. De er store i diameter. Bihulene i hodet er kjent som karoten sinus. Det venøse blodet fra karoten sinus kommer inn i kardinalhjertet.
Kardinalhjertet er blottet for muskler, og dets pumpevirkning er ikke på grunn av egenart, men skyldes sammentrekning av skjelettmuskler som omslutter den. Blodet fra kardinalhjertet mottas av anterior og posterior kardinalår. Den fremre kardinalvenen springer blodet direkte med grenhjerte mens den bakre kardinalvenen åpner seg inn i portalhjertet.
Det venøse blodet fra hud, kroppsmuskulatur og slimkjertler samles og helles i bakre kardinalvein. Den bakre kardinalvenen mottar også blod fra nyre og haleområde.
Etter å ha mottatt blod fra tarm, gonad, nyre og kroppsvegg og haleområde, åpner den bakre kardinalen inn i grenhjerte, dvs. sinus venosus ved separat åpning. Blodet fra subkutane bihuler kommer inn i kaudalt hjerte som henter sitt venøse blod i de caudale årene, mens det i sin tur åpner seg for kaudalvenen.
Fysiologi og innervering av grenhjerte i hagfisk:
Det er en stor kontrovers om hjertet av myksin er aneural (uten nerve) eller forsynt med nerver. Greene (1902) foreslo at hjertet av California hagfish er fri for all nervøs regulering og derfor må være aneural.
Greene (1902) konkluderte med at han ikke var i stand til å endre hastigheten og kraften i hjerteslag ved elektrisk stimulering av vagus nerver og av forskjellige andre deler av sentralnervesystemet.
Augustinson et al., (1956) rapporterte lignende resultater i hagfishen med vagal stimulering. Hirsch et al, (1964 og 1970) rapporterte at hjertet av Eptatretus stouti er forsynt med store nerver med myelinerte fibre og ganglioniske celler. De fant også ganglionceller med fibre i aorta, epikardium og endokardium.
De rapporterte også tilstedeværelsen av høyt katekolamininnhold i hjertevevene i California hagfish. Yamauchi (1980) bekreftet nærvær av nerver og nerveceller i hjertet av Eptatretus burgeri ved elektronmikroskopi. Ifølge ham viste neuronal perikaryon det fine strukturelle utseendet ganske typisk for nevronene i de høyere vertebrater.
Det er derfor klart nå at hjertet av hagfish ikke er aneural. Men på grunnlag av elektronmikroskopi holdt Yamauchi (1977) at ganglionceller langs den endokardiale kanten av hovedkanalen i ventrikelen beskrevet av Hirsch (1970), er bare catecholamin-lagring, spesifikke interstitiale celler.
Kardialcellene består av aktin og myosinproteiner, og på grunn av tilstedeværelsen av lignende bånd, er sammentrekningen lik skelettmuskulaturen.
Sinus venosus inneholder muskelceller. De spenner fra 9-15 μ diameter i bredde og har godt utviklet sarkoplasmisk retikulum med perifer kobling. Elektronmikroskopien viser at myofibrilene har A- og I-bånd med H- og Z-linjer i en sarkomerer-M-linje er også tydelig synlig i myofibrillen.
8. Reproduktive System av Hagfish:
Det er en enkelt gonad i hagfish. Det er tilstede på høyre side av tarmen som henger i bukhulen gjennom en delikat membran. Den strekker seg fra leveren og når opp til cloaca. Gonaden av hagfish er merkelig. Det er protandrisk hermafroditt som er i de tidlige stadiene det er mannlig og utvikler seg til kvinner.
I svært tidlige stadier av utvikling er gonadens vev neutralt, men inneholder elementene i begge kjønn, og det er vanskelig å finne ut om individet skal utvikle seg til mann eller kvinne. Det antas at i tidliglivet har den fremre regionen en moden eggstokk, mens den bakre regionen inneholder umodent testikkelvev.
Men når dyret forfaller, utvikler den mannlige og kvinnelige delen av gonaden enten til mann eller kvinne, og den andre delen blir undertrykt antagelig under påvirkning av hormoner fra hypofysen.
Gjødslingen er ekstern, ettersom kopulerende organer er fraværende i begge kjønn. Eggene er veldig store, keratinert med kroker. De er 2 til 3 cm i diameter. Eggene er forenet i begge ender av tuft av filamenter (figur 2.7a, b).

Prosessen hvor spermene kommer inn i egget for befruktning, er ikke klart forstått. Det tar mange måneder å utvikle eggene. Den hagfish legger vanligvis ned rundt 22 egg om gangen. Når de unge klekker seg fra egg, er de egentlig miniatyr voksne, så de undergår ikke den dramatiske metamorfosen som lamprey.