Forstå prosessen med celledivisjon (forklart med diagram)
Les denne artikkelen for å lære om prosessen med celledivisjonen (forklart med diagram)!
Eksistensen og levetiden avhenger av cellers funksjon og deres splittelse. Det er to viktige prosesser for celledeling, som er uløselig forbundet med hverandre for å opprettholde fremdriften i livet. Disse prosessene er mykose og meiosis.
Etter befruktning, når en sædcelle og en eggcelle forener for å danne en ny organisme, forventes det å bære dobbelt antall kromosomer fordi begge foreldrene bidrar med hele sitt antall kromosomer som passende for de respektive artene. Men prosessen meiosis forhindrer fordobling av kromosomene.
Det reduserer totalt antall kromosomer til halvparten, slik at avkom (nybildet celle) kan beholde det samme antall kromosomer som lik foreldre. På den annen side letter mitosen å danne to datterceller fra en enkelt celle hvor hver ny celle ligner foreldrecellen i alle henseender. I motsetning til meiose involverer mitose duplisering av kromosomale par, noe som er essensielt for cellegenerasjon, spesielt under veksten.
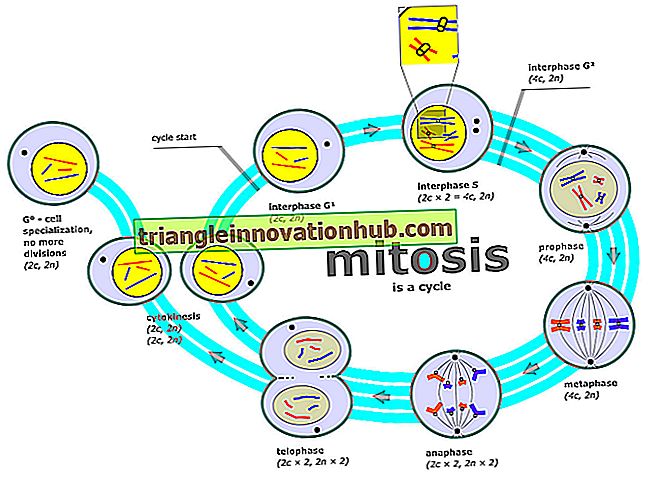
Mitosis mekanisme:
En spesifikk prosess kreves for å danne to datterceller ut av en modercelle. Denne prosessen er kjent som mitose og er vanligvis delt inn i fire stadier, profase, metafase, anafase og telofase. I noen tilfeller er det hensiktsmessig å betegne overgangen fra profase til metafase ved navn premetafase. Imidlertid er hele prosessen i syklusen av mitose en kontinuerlig en.

1. Profase:
Ved begynnelsen av profasefasen forbereder cellen seg for å dele kromosomene. Det hviler scenen når kjernen blir litt større. Det innebærer også kondensering av de tidligere diffuse, usynlige eller dispergerte trådene i synlige kromosomer.
Hvert kromosom er optisk dobbelt, som består av to identiske og tett parallelle tråder eller kromatider som ligger over hele deres lengde. Et kromosom bærer en innsnevring eller en smal region av festet til spindelen, som kalles som centromere.
Forskjellige andre enn sentromere kalles noen ganger 'sekundære sammenbrudd'. På profase-scenen ser centromere og sekundære innsnevringer ut, men ved metafase er dette skillet klart fordi de utvikler helt forskjellige relasjoner til spindelen.
I dette stadiet (profase) presenterer kretsidene en litt uregelmessig ull eller hårete utseende. De viser ikke generelt en serie granulater (krommer) som ses på den meotiske profasen, og sannsynligvis er dette en reell forskjell mellom de to. Ved profasens slutt forsvinner det ullete utseende nesten, og en jevn omriss er synlig.
2. Premetafase:
På slutten av profasen forsvinner atomvemnet vanligvis og dette trinnet kan defineres som perioden for spindeldannelse. I løpet av dette stadiet gir kromosomene et inntrykk av å slite og presse seg hverandre for å nå ekvator i utviklingsspindelen. Spindelen er relativt en solid gelatinøs kropp. Tilsynelatende består den nesten helt av protein med en svært liten mengde RNA.
Centrosen ser ut til å fungere som arrangør av spindelen. Centrosomet er den differensierte regionen av cytoplasma som inneholder centriol, som er et minutt granulat til stede like utenfor kjernemembranen. Etter at atomkraftmembranen er forsvunnet, fordobles centriolen og de to sentriolene beveger seg fra hverandre for å danne spindelens poler.
Spindelfibrene er trolig bunter av proteinmolekyler. Det kan skelnes mellom kontinuerlige fibre og kromosomale fibre. De kontinuerlige fiberene løper fra pol til pol, mens de kromosomale fibre forbinder sentromerer til polene.
I tilfelle av bipolare spindler er ekvatorialplanet plassert vinkelrett på lengdeaksen, som ligger midt mellom polene. Etter hvert som premetafasen fortsetter, ordner kromosomene seg med sine sentromerer, på ekvatorialplanet til spindelen. Det er den mest dynamiske scenen som gir opphav til en relativt statisk metafase.
3. Metafase:
Nukleolus forsvinner ved premetafasen og ved slutten av premetafasen viser kromosomene maksimal kondensgrad. Ikke bare det, på dette tidspunktet får kromosomene jevne utsnitt. De to kromatidene av hvert kromosom ligger parallelt eller løst viklet rundt hverandre, og forblir bare forenet ved sentromeren.
De ser nesten ut som delte kromosomer, men uten adskillelse av datterens sentromerer. Således er metafase et stadium hvor lite eller ingen synlig forandring finner sted i cellen; det er en statisk fase og vanligvis en kort en.
4. Anafase:
Anafasen er preget av begynnelsen på en separasjon av datter-sentromeren fra hverandre. Sentromeren splittes plutselig og deres halvdeler avviser aktivt hverandre. Da datterens sentromerer beveger seg opp mot spindelen mot motsatt poler, drar de kromatidene som er festet til dem, vekk fra hverandre.
Faktisk beveger de to halvdelene eller de samme settene av kromatider seg til to motstående poler av spindelen. Denne bevegelsen er autonom. Dermed starter den mitotiske separasjonen av kromatider alltid ved sentromeren og fortsetter langs kromosomene i begge retninger. Etter en fullstendig separasjon av kromatidene fra hverandre, kalles de datterkromosomer.
På dette tidspunktet blir spolens poler lenger enn hverandre i metafasespindelen. Datters centromerer blir også skilt fra hverandre og ekvatorialområdet mellom dem ser ut til å være lengre.
Når ekvatorialdelen av spindelen forlenger betydelig, blir polene tilsvarende innsnevret og til slutt ser det hele ut som en tynn bunke av trådlignende strukturer. Denne trådlignende strukturen mellom de to separerende gruppene av datterkromosomer kalles stamkroppen.

5. Telofase:
Som regel er de to gruppene av kromatider nå kjent som de to gruppene av datterkromosomer som passerer mot motstående poler av spindelen. Spindelens polarhoder ser ut til å forsvinne i begynnelsen av Telophase, men stamkroppen kan vare lenge.
De to gruppene av datterkromosomer mister raskt sine glatte konturer og gjennomgår de-kondensering. En tilsynelatende ny kjernefysisk membran som omkranser kromosomet utvikler seg på dette stadiet og nukleolus dukker opp igjen. Men i dette telofasetrinnet består kromosomet av bare ett kromatid i stedet for to, og det er bare halvparten av DNA-mengden, slik at utseendet av profase- og telofase-kjerner i samme vev nesten alltid er karakteristisk forskjellig. Imidlertid dannes to datterceller til syvende og sist.
Mitotisk cellefordeling er svært viktig fordi kromosomens bestanddeler aldri forandres og forblir identiske i alle dattercellene. Dermed er kjernen i kjærlighet lik moderkjernen. Som et resultat produserer prosessen med celledeling store antall datterceller, som gradvis begynner å anta forskjellige funksjoner, og på den måten blir de forskjellige kroppsdelene dannet.
Mekanismen for mesiose:
Kunnskap om arvelighet er viktig i måten å forstå mekanismen for meiosi. Faktisk er meiosis antitese av befruktning. I denne prosessen med celledeling blir det diploide antallet kromosomer redusert til haploid (halv) tallet. Det er en svært viktig prosess.
Siden vi vet at livet oppstår som en forekomst av fusjon mellom en mannlig og kvinnelig gamet, forventes antall kromosomer å være dobbelt fordi begge gametene bidrar med sitt spesifikke antall kromosomer.
Således vil kromosom nummeret fortsette å formere seg fra generasjon til generasjon. Naturens løsning på dette problemet er megiose eller reduksjonsdelingen. I høyere dyr foregår denne prosessen like før bakteriene blir dannet.
Meiosis av mannlige gameter blir referert til som spermatogenese mens samme mekanisme i kvinnelige gameter betegnes som oogenese. Imidlertid involverer meiosis to typer divisjoner, som forekommer etter hverandre. Ved første divisjon reduseres antall kromosomer til halvparten, men i andre divisjon blir nummeret dobbelt.
Meiosis er derfor en svært komplisert mekanisme for atomavdeling, som kan diskuteres på følgende måte:
Første Meiotiske Divisjon:
Den første "meotiske divisjonen" begynner alltid med et lengre profasestadium, som i stor grad er forskjellig fra en enkel mitotisk deling. Denne "Profeten" viser en rekke delstadier, som er kjent som leptotene, zygotene, pachytene, diplotene og diakinesis. Ved siden av Prophase er det en kort pre-metafase, som etterfølges av metafasen til den første 'meotiske divisjonen'. Vi skal imidlertid begynne med underfasen av Profet.
1. Leptotene:
Det er vanligvis en kort scene når kromosomene ser veldig langstrakte og slanke ut. Disse kromosomene har en polarisert orientering med alle sine ender rettet mot et lite område på den ene siden av kjernen. Denne polariseringen synes ikke å påvirke sentromerer i det hele tatt. Hvert kromosom bærer et bestemt antall granuler, som danner en kjede av perler av forskjellige størrelser kjent som kromomerene.
2. Zygotene:
I dette stadiet kommer de homologe kromosomene sammen i par og blir mer tette gjennom hele lengden. Dette fenomenet kalles paring eller synapsis. Denne synapsisprosessen virker som regel å starte ved kromosomendene og fortsetter deretter til den er fullstendig, bortsett fra noen kjønnskromosomer. Parring eller synaptisk prosess er utvilsomt tilrettelagt av den polariserte orientering av kromosomene, som bringer deres ender tett sammen.
3. Pachytene:
Pachytene begynner når synapsis kommer til en slutt, og det ender når de homologe kromosomene begynner å bevege seg vekk fra hverandre.
Hovedhendelsene som oppstår i denne fasen er som følger:
a) Det er ytterligere kondensasjon av kromosomer slik at kromosompar blir kortere og tykkere,
b) Som følge av synapsis mellom homologe kromosomer er antall observerbare kromosomer bare halvparten (dvs. n) av det somatiske kromosomnummeret. I de fleste av artene kalles disse haploide tallene av kromosompar bivalenter. Hver toverdige viser fire kromatider eller tråder. Disse strengene er omtrent like langt fra hverandre, slik at de danner et tverrsnitt i tverrsnitt.
c) Kryssing mellom homologe kromosomer foregår i løpet av dette stadiet. En liten mengde (0, 3%) DNA-syntese som finner sted under pachytene antas å spille en avgjørende rolle i krysset over. Pachytene er imidlertid vanligvis et langt stadium.
4. Diplotene:
Den synaptiske attraksjonen mellom de homologe kromosomene kommer plutselig til ende, og homologe kromosomer av hver bivalent begynner å bevege seg vekk fra hverandre. Dette markerer begynnelsen på Diplotene scenen. De to homologene til hver bivalent ser ut til å være festet med hverandre på ett eller flere punkter, kjent som Chaisma (flertall: Chiasmata). Det antas at i utgangspunktet ligger chiasmata ved punktene for faktisk kryssing mellom de homologe kromosomer.
Når diplotene utvikler seg, beveger chiasmata sakte mot endene av de homologe kromosomene. Denne bevegelsen er referert til som chiasma-terminalisering. Videre kondensasjon av kromosomer er også synlig i dette stadiet, slik at de ble gradvis kortere og tykkere.
5. Diakinesis:
Det er ingen vesentlig forskjell mellom sen diplotene og diakinesis. Faktisk vender de slanke diplotene bivalenter seg gradvis til tykke kromosomer og blir svært korte. De forblir spredt fordelt i hele kjernen, og alle sløyfer mellom chiasmata ligger i rette vinkler til hverandre. 'Rotasjon' er vanligvis fullført ved begynnelsen av diakinesis, men 'terminalisering' kan fortsette helt opp til første metafase.
Premetaphase:
I dette stadiet forsvinner atommembranen og spindelen dannes. Nukleolus er også funnet å forsvinne, og de bivalente kromosomene beveger seg mot ekvatorialplanet mens de to sentromerer av hver toverdige går på motsatte sider av flyet.
1. Første metafase:
I dette stadiet knytter kromosomene ofte til spindelen et sted mellom ekvator og polen. Sentromeren er festet til spindelfibrene på ekvator.
2. Første anafase:
Kromosomene blir nå skilt og beveger seg mot motsatt poler. Sentromererene deles ikke, men hver hel sentromere beveger seg i retning av nærmeste pol som dattercentromerer av en vanlig mitotisk deling.
3. Første telofase:
Denne scenen avviger ikke vesentlig fra det for en vanlig somatisk mitose. Her dannes to datterkjerner med et par kromatider ved polene. På denne måten inneholder to datterkjerner haploid antall kromosomer.
Andre Meiotiske Divisjon:
1. Second Prophase:
Den presenterer et tydelig stadium, som alltid er kort og inkluderer ingen komplikasjoner. Spindlene blir organisert raskt.
2. Andre metafase:
Antallet av kromosomer er halvparten av det somatiske tallet, og kromatidene avviger i stor grad, idet de bare holdes sammen ved sentromererne.
3. Andre anafase:
Sentromererene deler seg og begynner å bevege seg mot polene, siden kromatidene er ganske tre fra hverandre.
4. Andre Telophase:
Denne scenen er ikke forskjellig fra den somatiske delingen, unntatt antall kromosomer. Fire datterkjerner dannes, hver med haploid antall kromosomer. Det kan konkluderes med at den første meiotiske delingen forårsaker reduksjonen i antall kromosomer, men sentromeren mislykkes i å dele seg. Under den andre meiotiske delingen når sentromerer deler, deler ikke kromatidene seg.
De beveger seg ganske enkelt fra hverandre og danner fire datterceller av haploidnummer. Hovedprosessen av denne prosessen ligger i opprettholdelsen av et konstant antall kromosomer i arten. I alle seksuelt reproduserende planter og dyr er gametene haploid (X) i antall kromosomer. Seksuell reproduksjon er sammensmeltingen av en mannlig gamete (X) med en kvinnelig en (X) og resulterer i dannelsen av en zygot (2X) hvorfra et avkom utvikles. En annen betydning av meiosis er å hjelpe segregering, assortiment og rekombination av gen.